Liebe Ditte und alle anderen Risspilz-Interessierten,
ein Inocybe aus China ist mir noch nicht untergekommen. Besten Dank für das Portrait.
Die Sporenform ließen zunächst Zweifel aufkommen. Visuell kamen sie mir etwas breit vor. Deine angegeben Maße passen aber perfekt ins Konzept.
Wie von Dir schon erwähnt steht I. heimii (= I. caesariata) sehr nahe. Die artspezifischen Merkmale lassen sich wie folgt kurz zusammen fassen: Hut kräftig wollig schuppig; Stiel kürzer als Hut breit und gewöhnlich mit verjüngter Basis, Cortina weißlich und kräftig ausgeprägt, nicht selten dauerhaft mit ringartigen Velumresten am Stiel verbleibend; Sporen schlank, Qm aber nicht deutlich über 2 gehend, Lamellenschneide nicht homomorph, Cheilozystiden schlank keulig.
Zu I. malenconi ein Auszug aus einer noch nicht publizierten Arbeit über die gesamte „Dulcamara-Gruppe“:
Inocybe malenconi Heim, Wollfaseriger Risspilz
Beschreibung eines Fundes aus dem NSG Zechau im Altenburger Land (Thüringen) bei Meuselwitz:
Hut bis 28 mm breit, flach konvex, später niedergedrückt, Rand kaum abgebogen; ockergelb, ockerbraun bis orangebraun, alt dunkel rotbraun, teils mit hellerer Randzone; filzig bis schuppig-, stellenweise und vor allem im Alter auch fast glatt.
Lamellen normal gedrängt, ca. 1:1 untermischt, gerade, breit bis etwas herab laufend am Stiel angewachsen; lange hell bleibend, im Alter ocker ohne olivfarbene Töne; Schneide heller, feinst bewimpert.
Stiel bis 35 x 4 mm, zylindrisch, Basis teils mit angedeutetem kleinem Knöllchen, meist aber nicht verdickt; auf bräunlichem Grund weißlich, anliegend überfasert, Spitze flockig.
Cortina weißlich, flüchtig.
Trama weißlich-bräunlich; Geruch wie bei I. dulcamara mit honigartiger Komponente.
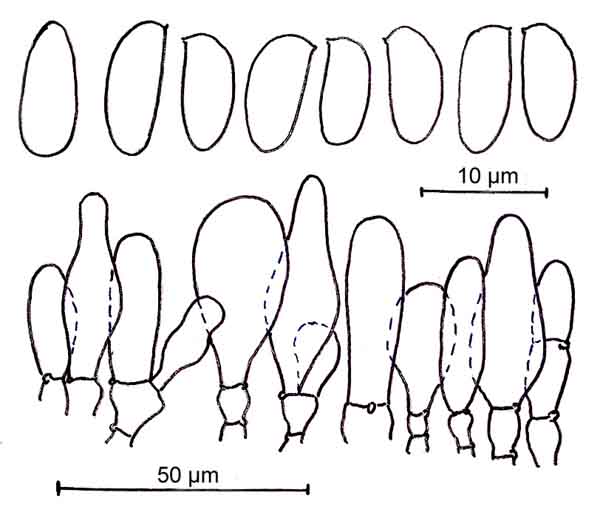
Sporen 8,3-10,6 (11) x (3,8) 4,1-4,9 (5,2) µm (im Mittel 9,75 x 4,5 µm), Q (1,8) 1,9-2,35 (2,55) (Qm 2,15), Vm 110 µmł; zylindrisch bis schlank bohnenförmig, nur ausnahmsweise schwach laceroid, apikal vorwiegend abgerundet.
Basidien 4-sporig, keulig, 22-38 x 6-10 µm; farblos.
Cheilozystiden gedrängt stehend; teils kettenartig gegliedert, Endglieder keulig bis zylindrisch, seltener eiförmig, subutri- und subfusiform, 15-50 (58) x 8-17 (20) µm; farblos und dünnwandig.
Stielbedeckung nicht näher untersucht. HDS nicht untersucht.
Funddaten: 03. X. 2003, Thüringen, Altenburger Land bei Meuselwitz, NSG Zechau (ehemaliger Braunkohlentagebau), MTB 4939/4, ca. 200 m ü. NN, sehr feuchte, von kleinen Quellen umgebene Lage unter Populus balsamifera (Balsampappel); Beleg AV031004a
Im Gegensatz zur Varietät megalospora STANGL & BRESINSKY sind dem Verfasser von der Typusvarietät nur noch zwei weitere Fundmeldungen aus Deutschland bekannt:
Ein sächsischer Fund liegt zeitlich wie auch räumlich kaum vom oben beschriebenen entfernt und auch das Habitat war, da es sich ebenfalls um eine Bergbaufolgelandschaft handelte, wohl kaum wirklich verschieden. Auch waren bei dieser Kollektion die Hüte im jungen Zustand relativ hell ockergelblich bis ocker, was in etwa der Abbildung bei ALESSIO & REBAUDENGO (1980) entspricht, später aber braun bis rot- und deutlich dunkler, was mehr der Abbildung bei HEIM (1930) nahe kommt. Bei Nachfolgefunden vom selben Ort konnte dieses Nachdunkeln mit zunehmendem Alter der Fruchtkörper ebenfalls beobachtet werden (T. RÖDEL, mündliche Mitteilung), so dass die Hutfarbe nur bedingt als aussagekräftiges Kriterium geeignet sein kann. Die mikroskopischen Unterschiede zur oben beschriebenen Kollektion waren sehr gering und beschränkten sich zum einem auf die Sporen, die bei gleicher Länge geringfügig schmaler waren und zum anderem auf die Cheilozystiden, die durch eine ziemlich variable Form auffielen.
06. X. 2003, Sachsen, Hochhalde Trages bei einem früheren Braunkohlentagebau bei Espenhain, MTB 4841/1 unter Populus tremula und Salix spec.; leg. & det. T. RÖDEL.
Ein zweiter thüringer Fund vom Nordrand des Thüringer Waldes, könnte auf Grund der bei mehreren Basidiocarpien beobachteten auffallend gelben Stielbasis genau so gut innerhalb des Fulvipes-Komplexes eingeordnet werden. Zur Kenntnis der Variationsbreite der seltenen Art, wird auch von dieser, aus anderem Habitat stammender Aufsammlung eine kurze Beschreibung gegeben.
28. V. 2006, Thüringen, Ilmenau, MTB 5331/1, ca. 450 m ü. NN, unter Corylus, Betula und Tilia; leg.: F. HAMPE, det. A. VESPER; Beleg AV060528a.
Hut Ř bis 50 mm, konvex, später flach konvex, im Zentrum angedeutet stumpf gebuckelt bis schwach genabelt, Rand wenig abgebogen, scharf; gelbocker bis orangebraun mit wenig hellerer Randzone; am Scheitel matt, teils grindig aufgerissen, nach außen kurz radialfaserig bis etwas faserschuppig. Lamellen ziemlich dicht stehend, 1:1 bis 1:3 untermischt, bis 8 mm breit und in voller Breite am Stiel angewachsen; beige, rosabräunlich bis rotbraun mit hellerer, bewimperter Schneide. Stiel bis 30 x 8 mm, zur Basis erheblich verjüngt; apikal blass gelblich und flockig, darunter überfasert, teils auch nach oben abstehend faserschuppig und ähnlich dem Hut gefärbt, 1x mit angedeuteter faseriger Ringzone; meist enghohl; Basis weiß filzig, an verletzten Stellen auffallend gelb.
Cortina nur rudimentär erhalten, blass ocker.
Trama im Hut weißlich, im Stiel schmutzig weißlich bis bräunlich, im Innenbereich der Stielbasis bei einzelnen Fruchtkörpern teils auffallend gelb; Geruch erdig, im Schnitt auch mit leichter honigartigen Komponente.
Sporen 8,4-11,0 x 4,2-5,1 µm (im Mittel 9,5 x 4,5 µm), Q (1,8) 1,9-2,3 (2,4) (Qm 2,1), Vm 105 µmł; zylindrisch bis schlank bohnenförmig, nur ausnahmsweise laceroid, apikal vorwiegend abgerundet.
Basidien 4-sporig, keulig, 28-42 x 7-9 µm.
Cheilozystiden vielgestaltig, Endglieder breit keulig, lageni- bis fusiform, einige auch subutriform und angedeutet kopfig, 23-45 x 9-22 µm; farblos; Wand bis 1 µm dick; vereinzelt mit Basidien vermischt.
Hyphen der Stielbedeckung gelblich, glatt bis angelagert inkrustiert; im oberen Bereich mit einigen kettenförmig gegliederten kaulozystiden Elementen vermischt.
Tramahyphen der Stielbasis fast von einer „Dryophila-Struktur“; blass bis kräftig intrazellulär gelblich pigmentiert.
HDS aus schlanken, wenig inflaten, ca. 25-100 x 6-13 (16) µm großen, gelbbraunen, nur ausnahmsweise angelagert inkrustierten Hyphen; Wandstärke bis 1 µm.
Diese Kollektion passt zu keiner der von KÜHNER mit gelber Stielbasis beschriebenen Pilze (siehe Fulvipes-Komplex), wohl aber auf Grund der meisten makroskopischen, hauptsächlich aber der mikroskopischen Merkmale, gut zur schlanksporigen Inocybe malenconi. Auffallend war die große Variabilität der Cheilozystiden, die ähnlich so beim sächsischen Fund zu beobachten war.
Erstmals wurde dieser Risspilz 1928 von MALENÇON in Frankreich im Großraum von Paris auf feuchtem Grund gefunden und von HEIM (1931), leider nur knapp, beschrieben. Spätere Fundmeldungen sind nur spärlich in der Literatur zu finden. So berichten STRIDVALL & JACOBSSON (1989) von schwedischen Funden bei Salix und geben als Schlüsselmerkmal neben den schlanken Sporen mehr rundliche Cheilozystiden an. Bei HEIM (1931) finden sich keine Angaben dazu und BON (1997) beschreibt sie unauffällig keulig, gibt aber nicht an, auf welche Funde oder Quellen er sich bei seiner Kurzbeschreibung bezieht. Dagegen werden die Cheilozystiden der Japanischen Kollektionen von KOBAYASHI (2002) als nahezu zylindrisch genannt, gezeichnet sind sie aber deutlich keulig! Die Sporenmaße der Europäischen Funde unterscheiden sich nur minimal (Tab. 1). Anders wieder bei der Japanischen Aussammlung, wo diese breiter und in Form auffallend elliptisch angegeben bzw. gezeichnet sind (KOBAYASHI 2002), so dass diese Aufsammlungen nicht mit Sicherheit dieser Spezies zugerechnet werden kann.
HEIM 9,0 - 12,0 x 4,0 - 4,8 µm
STRIDVALL & JACOBSSON 10,0 - 12,0 x 4,0 - 5,0 µm
BON 9,0 - 12,0 x 4,0 - 5,0 µm
KOBAYASHI 9,7 - 11,7 x 4,1 - 6,3 µm
RÖDEL 8,0 - 11,0 x 3,8 - 4,9 µm
VESPER 8,3 - 10,6 x 4,1 - 4,9 µm
HAMPE 8,4 - 11,0 x 4,2 - 5,0 µm
Die Differenzierung zur großsporigen Sippe I. megalospora beschränkt sich im Wesentlichen auf die signifikant größeren Sporen, bei weitestgehend identischer Sporenform. Im Verbreitungsatlas der Großpilze Deutschlands (KRIEGLSTEINER 1991) wurde diese Varietät nicht aufgenommen. Wie aber Fundmeldungen in Floren und an den Verfasser belegen, scheint diese Varietät in einigen Regionen Deutschlands nicht ausgesprochen selten zu sein. So z.B. in Sachsen, unweit von Thüringens Grenzen bei Zwickau (F. FISCHER) und im Leipziger Land (T. RÖDEL), sowie in Bayern, wo 3 ältere Fundmeldungen von J. STANGL und A. EINHELLINGER (STANGL 1989) und A. BRESINSKY (ZITZMANN 2002) vorliegen. Die Angabe bei KRIEGLSTEINER (1999) aus dem Naturraum Mainfränkische Platten mit Sporenangaben (10)-12-14-16,5 x (6)-7-7,5-(8,5) µm und einem Sporenquotienten unter zwei kann kritisch bewertet nicht dieser Art zugerechnet werden und lässt eher verwandtschaftliche Nähe zum Formenkreis um Inocybe fuscomarginata erahnen. Eine verlässliche Aussage zur Ökologie der beiden Varietäten ist bei den relativ wenigen Nachweisen kaum möglich. Vermutlich bevorzugt aber die Typusvarietät eher schattige, feuchte Lagen, die Varietät megalospora scheint dagegen vorzugsweise über trockenen, silikatreichen und nicht zu sauren Böden zu gedeihen.
Verwendete Literatur (eine Auswahl zu den hier erwähnten Arten habe ich nicht erst noch getroffen):
ALESSIO, C. L. & E. REBAUDENGO (1980): Inocybe-Supplementum III (Vol. XXIX) zu G. BRESADOLA, Iconographia Mycologica. Trento.
ARNOLDS, E., T.W. KUYPER & M.E. NOORDELOOS (Hrsg.) (1995): Overzicht van de paddestoelen in Nederland. Beilen.
BIZIO E. (1995): Alcune Inocybe piů frequentidella zona alpinadelle Dolomiti.- R.d.M. (AMB) Suppl. n. 2: 3-60.
BIZIO E. & E. FERRARI (1999): Il genere Inocybe (Fr.) Fr. nel Verbano-Cusio-Ossola e in alcune zone limitrofe.- Funghi e Ambiente 80-81: 13-82.
BOLLMANN, A., A. GMINDER & P. REIL (2002): Abbildungsverzeichnis mitteleuropäischer Großpilze. Hornberg.
BON, M. (1988): Pareys Buch der Pilze. Hamburg, Berlin.
BON, M. (1997) : Clé monographique du genre Inocybe.- Doc. Myc. 27 (105): 1-51.
BREITENBACH, J. & F. KRÄNZLIN (2000): Pilze der Schweiz. Bd. 5. Luzern.
DÖRFELT (Hrsg.) (1988): BI-Lexikon Mykologie-Pilzkunde. Leipzig.
EBERT, P. (1960): Inocybe dulcamara (ALB. et SCHW. ex FR.) QUÉL. als Aschepilz.- Westfälische Pilzbriefe Bd. 2: S. 92-96.
EINHELLINGER, A. (1987): Die Gattung Russula in Bayern. Berlin, Stuttgart.
FERARRI, E. (2006): Inocybe alpine e subalpine.- Funghi non delineati. Pars XXXIV-XXXVI.
HARDTKE, H.-J. & P. OTTO (1999) Rote Liste Pilze Freistaat Sachsen. Dresden.
HEIM, R. (1931): Le Genre Inocybe. Encyclopédie mycologique 1. Paris.
HORAK, E. (2005): Röhrlinge und Blätterpilze in Europa. München.
JAMONI, P. G. (2000): Le Inocybe della zona alpina del Monte Rosa Vercellese.- Funghi e Ambiente 82: 3-60.
KAJAN, E. (1988): Pilzkundliches Lexikon. Schwäbisch Gmünd.
KOBAYASHI, T. (2002): The taxanomic studies of the genus Inocybe.- Nova Hedwigia, Beiheft 124: 1-368. Stuttgart.
KREISEL, H. (Hrsg.) (1987): Pilzflora der Deutschen Demokratischen Republik. Jena.
KRIEGLSTEINER, G. J. (1991a): Verbreitungsatlas der Großpilze Deutschlands (West), Band 1: Ständerpilze, Teil B: Blätterpilze. Stuttgart.
KRIEGLSTEINER, G. J. (1991b): Über neue, seltene, kritische Makromyzeten in Westdeutschland.- Beiträge zur Kenntnis der Pilze Mitteleuropas VII: 61-79.
KRIEGLSTEINER, L. (1999): Pilze im Naturraum Mainfränkischen Platten und ihre Einbindung in die Vegetation.- Regensburger Mykologische Schriften 9(1-2): 1-905.
KÜHNER, R. (1955): Compléments a al „Flore Analytique“ VI. Inocybe goniosporés et Inocybe acystidés. Espčces nouvelles ou critiques.- BSMF 71 (3) : 169-201.
KÜHNER, R. (1988): Diagnoses de Quelques nouveaux Inocybes récoltés en zone alpine de la Vanoise (Alpes françaises).- Documents mycologiques Tome XIX – Fascicule no. 74: 1-27.
KUYPER, T.W. (1986): A revision of the genus Inocybe in Europe.- Persoonia Suppl. Vol. 3: 1-247.
MALENCON G. & R. BERTAULT (1970): Flore de Champignons Supérieurs du Maroc. Tome I. Rabat.
MICHAEL, H., B. HENNING & H. KREISEL, H. (1981): Handbuch für Pilzfreunde. IV. Blätterpilze - Dunkelblättler. Jena.
MONTAG, K. (2000): Porträt Nr.34.- Der Tintling 24.
MONTAG, K. (2001): Leserforum.- Der Tintling 25: 6.
MOSER, M. (1983): Die Röhrlinge und Blätterpilze (Agaricales).- Kleine Krypt.flora II/b2, Basidiomyceten, II. Teil. Stuttgart.
MOSER, M. & W. JÜLICH (1985 ff.): Farbatlas der Basidiomyceten. Stuttgart.
NESPIAK A. (1990): Inocybe.- Grzyby (Mycota) Tom XIX: 1-175. Warszawa, Kraków.
NEUHOFF, W. (1960): Noch ein beachtenswertes Vorkommen des Risspilzes Inocybe dulcamara (ALB. et SCHW. ex FR.) QUÉL.- Westfälische Pilzbriefe Bd. 2: 122-126.
PEITNER, U. & al. (1999): Ergebnisse der 26. Mykologischen Dreiländertagung in Rotholz – Jenbach (Tirol) vom 29. 08. bis 05.09.1998.- Österreichische Zeitschrift für Pilzkunde 8, 83-124.
SCHMID-HECKEL, H. (1988): Pilze in den Berchtesgadener Alpen. Nationalpark Berchtesgaden, Forschungsbericht 15, S. 1-136, Regensburg.
STANGL, J. (1989): Die Gattung Inocybe in Bayern.- Hoppea 46, 1-394.
STRIDVALL L. u. A. & S. JACOBSSON (1989): Släktet Inocybe i Sverige.- Joordstjärnan 10 (2) 29-76.
WEHOLT, Ř. (1983): Notater om Inocybe.- Agarica 4, 283-291.
ZITZMANN, H. (2002): Die Gattung Inocybe (Agaricales, Cortinariaceae).- Regensburger Mykologische Schriften 10, 135-288.
Mikrozeichnungen: Sporen und Cheilozystiden von Kollektion RÖDEL
Herzliche Grüße
Andreas